


研究概要
これらの研究は、科学技術振興事業団・戦略的基礎研究事業「脳の神経回路形成と可塑性の分子機構」及び「細胞内パターニングによる組織構築メカニズム」、文部科学省、特定領域研究「細胞外環境ー神経ガイダンス制御分子CRMPによる細胞内・外位置情報伝達と組織構築」、ターゲットタンパク研究プログラム「セマフォリンおよびセマフォリン受容体分子群をターゲットにした構造・機能解析と治療法開発」、および種々の文部科学省科学研究費補助金の助成により行われています。
1.神経伝達物質ドーパの研究
新規の生体内活性物質の検索とその受容体の同定は生命科学の最も重要な課題であるだけでなく、創薬と臨床応用に結びつく可能性を秘めています。L-ドーパ(ドーパ)は、神経伝達物質ドパミンの前駆体であり、脳内ドパミンが枯渇して発症すると考えられているパーキンソン病の最も有効な薬物です。世界で最も権威のある薬理学教科書であるGoodman & Gilman's The Pharmacological Basis of Therapeuticsをはじめ、現在すべての教科書には、ドーパ自身には活性がないと記載されています(図1)。

1986 年以来、本研究室においては、世界に先駆けて、従来単なる神経伝達物質ドーパミンの前駆体と位置付けられてきたドーパが、それ自身で神経伝達物質ないし修飾物質として働くことを示してきました(Misu and Goshima, TiPS 14; 119-123, 1993; Misu et al, TiPS 23; 262-268, 2002)。この一連の研究は、ラット脳スライス灌流標本において電場刺激がドーパの遊離を引き起こすことを発見したことに始まります。「前駆体であるはずのドーパが遊離する」などと一体誰が考えつくでしょう。つまりこの仕事は生のデータを良く観察することの重要性を物語っています。もしドーパが伝達物質であるとするなら・・・・。先人たちの築いた伝達物質という概念に沿って検証を進めてきました。その結果、ドーパが従来知られている神経伝達物質と同様に電気刺激で遊離され、その遊離はナトリウムチャネル阻害剤のテトロドトキシン(フグ毒)で阻害され、外液からカルシウムイオンを除去すると殆ど起こらなくなることが判明しました。
ドーパは生体内で速やかにドーパミンへと酵素学的に変換されます。これはドーパがパーキンソン病という脳内ドーパミンが枯渇する病態に使用される根拠になっています。ドーパが伝達物質であるなら、ドーパミンへ変換せずとも一定の生物学的効果が出るはずです。実際、ドーパをドーパミンへと変換する芳香族L-アミノ酸脱炭酸酵素(AADC)を阻害しても、ドーパはドーパミンとは全く異なる効果を示すことが見出されました。
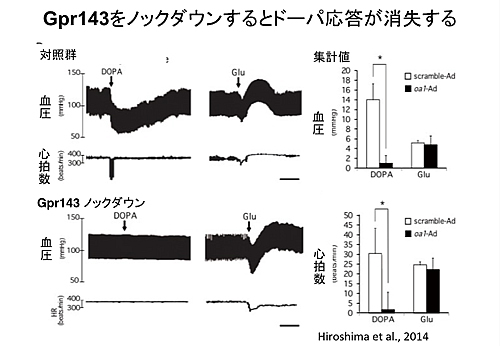
その後の一連の研究から、ドーパ拮抗薬の存在、神経線維の刺激による遊離と生理学的応答、拮抗薬による阻害、ドーパ含有ニューロンの存在など、ドーパが伝達物質共通の基本的な性質を備えていることの証拠をいろいろとを見出してきました(Neurobiology of DOPA as a neurotransmitter, ed. by Misu and Goshima, CRL Press, 2006)。つい最近になり、ドーパ受容体候補分子として長らくリガンドが不明であったオーファン型Gタンパク質連関型受容体 Gpr143 がドーパをリガンドとして認識することが明らかとなり、ドーパ神経伝達物質仮説の最終的な証明に向けて大きく一歩前進しました (Hiroshima et al., Brit J Pharmacol 171, 403-414, 2014)。この他にも、Gpr143が中枢神経系のみならず末梢臓器・神経系に広範に発現していること、今までに全く知られていなかった作用が続々と見つかるなど、私達の研究室のテーマの中で今最もホットな研究が展開しつつあります。薬理学の教科書が塗り替えられる日も間近と言えます。
2.細胞内軸索ガイダンス制御分子 CRMP の生物学的機能
神経突起伸展と特異的回路形成の過程には時間的・空間的に制御された様々な細胞-細胞間の認識機構、細胞内分子間の相互作用が存在すると考えられています。しかし、その分子機構の詳細は不明です。神経突起の伸展制御には大きく誘引と反発の2つの作用が関わる事が明らかになっています。私達は反発性分子であるセマフォリン(コラプシン)の情報を媒介する分子としてCRMP(Collapsin Response Mediator Protein)を同定しました(Goshima et al, Nature 376, 509-514, 1995)。同分子の作用機構はしばらく不明のままでしたが、 最近になり複数の研究室から報告が相次ぎ、多くのことが明らかになりつつあります。
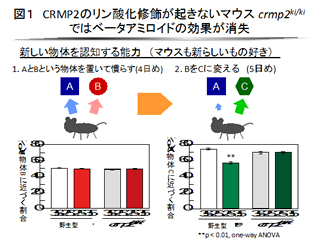
私達は線虫および哺乳類CRMPと相互作用する分子のスクリーニングを行っています。軸索伸長制御、スパイン成熟などのセマフォリン応答に関わる複数のリン酸化酵素を含む細胞内分子を新たに見出しました(Sasaki et al., Neuron 35; 907-920, 2002; Uchida et al., Genes Cells 10: 165-179, 2005)。また驚くべきことに、CRMPはリーリンという細胞移動、ひいては大脳皮質の形成に関わる細胞外分子のシグナルをも媒介することが最近になって明らかになりました(Yamashita et al., J Neurosci 26; 13357-13362, 2006)。CRMPと相互作用する分子や制御機構、細胞内局在に関わる分子機構、ならびにアルツハイマー病や統合失調症などの病態との関連性が今後の重要課題です。最近、アルツハイマー病の原因の一つとして重視されてきたβアミロイドという脳内タンパク質の毒性が、CRMP2のリン酸化修飾を介して発現することを見出しています (図1、2:Isono et al., Neurosci Res 77, 180-185, 2013)。
さらに文部科学省「科学技術振興調整費」に「翻訳後修飾プロテオミクス医療研究拠点の形成」という課題で採択され、私達の研究グループでは「修飾異常蛋白質の同定、バイオマーカーの開発」というテーマで研究を進めつつあります。具体的には、本研究室において機能解析を進めている複数の修飾蛋白質 (CRMPファミリー分子など)に着目しつつ、病理学、精神科や神経内科教室と共同して、病理組織検体やヒト血液検体を解析、これらの情報をもとに新規バイオマーカー候補を見いだすべく研究を進めています。こうした取組等を通じて臨床研究に有用なバイオマーカーの開発、さらには、本学生命医科学の構造生物学の研究グループと共同して、各疾患に選択的に作用する薬物の創成(創薬)へとつなげようとしています。
 |
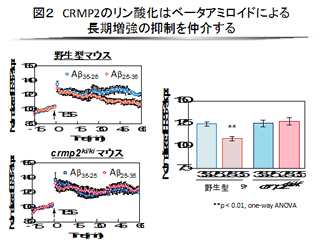 |
| 図1 ベータアミロイドの認知障害作用は、CRMP2のリン酸化修飾を介して起こる | 図2 ベータアミロイドの海馬における神経伝達障害は、CRMP2のリン酸化修飾を介して起こる (Isono et al, 2013) |
3.軸索ガイダンス分子Sema3Aによる細胞内輸送制御の生理学的役割
細胞が生体内で正常に働くためには細胞の方向性や細胞内外の特定の部位にある決まった分子が局在していることが必要と考えられるので軸索ガイダンス分子による細胞内輸送という現象には様々な制御機構や病態に重要な関わりがあると考えています。 神経ガイダンスという用語は、神経回路が正確に形成される過程における制御を示しています。従来、線虫やショウジョウバエの神経回路形成が異常となる変異体の解析を通じて、神経回路形成に関わる重要な分子が次々と同定されてきました。
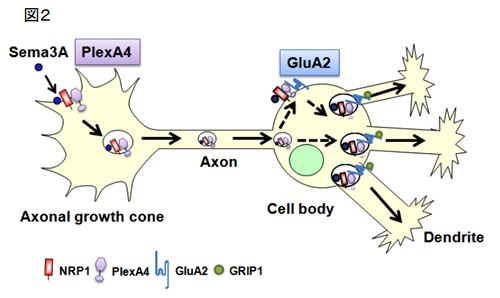
1997 年、私たちは、軸索ガイダンス分子のひとつであるSema3Aによって軸索内輸送axonal transportが亢進することを世界に先駆けて初めて報告しました(Goshima et al., J Neurobiol 33; 316-328, 1997)。この生理学的意義は長らく不明でしたが、Sema3Aによる成長円錐での局所蛋白合成にこの細胞内輸送が密接に関わる証拠を得ました(Li et al., J Neurosci 24; 6161-6170, 2004)。この論文は、Nature Neuroscience Review のハイライトに取り上げられました。 さらに、私達はSema3Aによる軸索輸送の生理学的意味を検討しました。まず、Sema3Aが一体、神経細胞のどこに効いて軸索輸送を引き起こすのかを微小ピペットを使って調べました。その結果、培養海馬神経細胞では、軸索先端の成長円錐局所に投与したときのみ、軸索輸送の促進が見られました。驚くべき事にこの時、軸索輸送の反応には少し遅れて、樹状突起における輸送も促進することがわかりました(図1)。
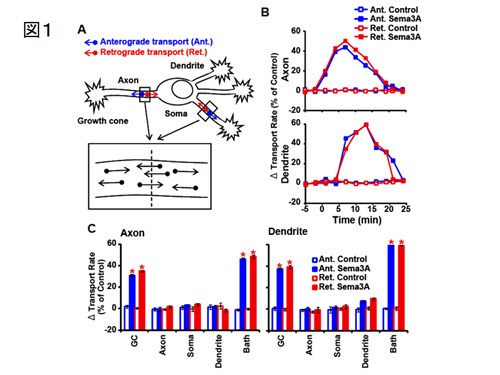
この結果は、軸索先端にある成長円錐から何らかのシグナルが伝達され、樹状突起に運ばれることを示唆します。この仕組みを調べるために様々な実験を行った結果、イオンチャネル (Yamane et al, BBRC 422, 333-338, 2012) と軸索輸送の双方が連関する全く新しい様式によって軸索先端から細胞体・樹状突起へとシグナルが伝達され、しかも、そのシグナルは結果として樹状突起の遠位端へのグルタミン酸受容体の輸送を促すことを発見しました(図2、Yamashita et al, Nat Commun, 2014)。
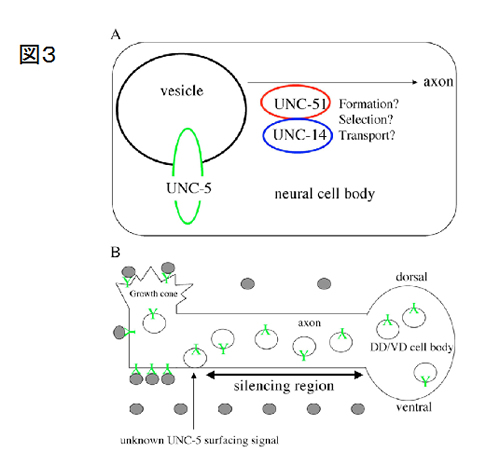
またモデル生物として線虫をとりあげ、軸索ガイド機構と細胞内輸送との関連を解析しています(図3、Ogura and Goshima, Development 2133; 3441-50, 2006)。unc-5変異体線虫では、DD/VDというニューロンの走行が異常になります。UNC-5は、UNC-6/ネトリンという軸索ガイダンス分子の受容体です。UNC-5が細胞外に分泌されたUNC-6/ネトリンの情報を受け取り、軸索は分泌源から遠ざかるように反発されます。もし、軸索ガイダンス分子やその受容体が生体内で正しい場所に位置することが出来ない場合、神経回路は正常に行われないと考えられます。
UNC-5の局在が異常となる変異体を探索したところ、unc-51変異体ではUNC-5の局在が異常となっていることを発見しました。 UNC-51は酵母のオートファジーに必要なAPG1に相同なセリン/スレオニンキナーゼであり、unc-51変異体ではDD/VDニューロンの軸索ガイドが異常になります。またunc-51の結合分子はRab, Rap等のGTPaseのシグナルに重要なRUNドメインを持っています。私達はプロテインフォスファターゼのPP2Aの触媒サブユニットのLET-92/PP2A-CがUNC-51と相互作用すること、let-92/PP2A-Cとunc-51が遺伝的に相互作用すること等を見出し、UNC-51によるリン酸化活性とPP2Aによる脱リン酸化活性のバランスが、軸索ガイダンスに大事な役割を演じている事を示しました(Ogura et al, Development 137, 1657-1667, 2010)(図3) 。
私達は他にもネトリン/UNC-6の局在が異常となる変異体を単離しています (Asakura et al, Dev Biol 304, 800-810, 2007)。 現在は、unc-51変異体が示す細胞極性異常という表現型に着目し、細胞極性と軸索・樹状突起パターン形成との連関機構について研究を進めています。
細胞が生体内で正常に働くためには細胞の方向性や細胞内外の特定の部位にある決まった分子が局在していることが必要と考えられるので軸索ガイダンス分子による細胞内輸送という現象には様々な制御機構や病態に重要な関わりがあると考えています。
4.軸索ガイダンスにおけるチロシンフォスファターゼの役割
私達は軸索ガイダンス分子のSema3Aによる成長円錐退縮反応において LAR クラスの チロシンフォスファターゼ が 関与するか否かを検討しています。軸索ガイダンス機構におけるチロシンフォスファターゼの関与は、ショウジョウバエにおける遺伝的解析から以前より指摘されていますが、そのメカニズムの詳細はほとんど手つかずの状態にあり、今後の進展が期待されます。