研究内容
カキの甘渋性制御機構
本研究室では,播種直後,開花結実が見られる前にカキの甘渋性を判別するための分子マーカーの開発を行っています.これまでに,甘ガキ育種の早期選抜に利用できる分子マーカーを開発しました.現在,このマーカーを利用して,甘渋性制御機構解明のための実験を行っています.また,日本の甘ガキとは異なる脱渋機構をもつ中国の甘ガキについて,その脱渋制御機構を解明するための研究を行っています.この中国の甘ガキを利用した甘ガキ育種も果樹研究所と共同研究として実施しています.

第1図 中国の羅田県で最近発見された甘ガキ
参考資料
- Kanzaki, S., T. Akagi, T. Masuko, M. Kimura, M. Yamada, A. Sato, N. Mitani, N. Utsunomiya, and K. Yonemori. 2010. SCAR markers for practical application of marker-assisted selection in persimmon (Diospyros kaki Thunb.) breeding. J. Japan. Soc. Hort. Sci. 79: 150-155.
- Akagi, T., R. Tao, T. Tsujimoto, A. Kono, and K. Yonemori. 2012. Fine genotyping of a highly polymorphic ASTRINGENCY-linked locus reveals variable hexasomic inheritance in persimmon (Diospyros kaki Thunb.) cultivars. Tree Genetics & Genomes 8:195-204.
自家不和合性制御機構
オウトウやアーモンド,ウメやスモモなどのサクラ属果樹には自己花粉が受粉しても受精できない自家不和合性という現象があり,栽培や育種の障壁となっています.本研究室では,不和合性認識メカニズムの解明と得られた知見の栽培・育種的利用に関する研究を行っています.これまでに自家不和合性雌ずい側因子であるS-RNaseと花粉側因子であるSFB (S haplotype-specific F-box protein gene)を同定しました.現在,自己・非自己認識の分子機構の解明を進めています.

第1図 花柱タンパク質の二次元電気泳動解析
参考資料
- Yamane, H. and R. Tao. 2009. Molecular basis of self-(in)compatibility and current status of S-genotyping in rosaceous fruit trees. J. Japan. Soc. Hort. Sci. 78: 137-157.
- Tao, R. and A.F. Iezzoni. 2010. The S-RNase-based gametophytic self-incompatibility system in Prunus exhibits distinct genetic and molecular features. Scientia Horticulturae 124: 423-433.
休眠制御機構
温帯果樹の多くは,冬季には活動を休止し休眠とよばれる状態で越冬します.開花には一定時間以上の低温遭遇が必要であり,開花時期決定には秋から春にかけての温度変化が影響しています.近年の気候変動により,低温不足による開花の不揃い,あるいは開花前進による霜害の多発など,果樹の栽培現場でいくつかの問題が生じています.本研究室では,温帯果樹の休眠の制御機構の解明とその人為制御法の開発を目指して研究を行っています.
参考資料
- Yamane, H. (2014) Regulation of bud dormancy and bud break in Japanese apricot (Prunus mume Siebold & Zucc.) and peach [Prunus persica (L.) Batsch]: A summary of recent studies. J. Japan. Soc. Hort. Sci. 83: 187-202.

第1図 亜熱帯地域で露地栽培されているニホンスモモの様子.
休眠覚醒に必要な低温要求量が十分に満たされず,同じ枝のなかで開花ステージが同調していない
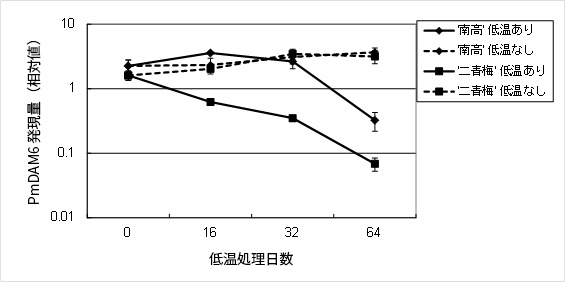
第2図 低温に応答する休眠制御遺伝子PmDAM6遺伝子の発現変化
‘南高’は低温64日目で‘二青梅’は低温32日目で明確な休眠覚醒が観察された.両品種とも低温にさらされない対照実験では休眠から覚醒しなかった.‘南高’では低温64日目に,‘二青梅’では低温16-32日目に,PmDAM6遺伝子発現量の低下がみられた.一方,非低温処理区では発現低下がみられなかった.現在,休眠を正に制御する低温応答性転写因子であるDAM遺伝子のシグナル伝達経路の解明を試みている.
雌雄性制御機構
「性」の決定は生物が進化の中で獲得した多様性の維持のための機構の一つであり, 作物の育種・栽培の両面において最大限に考慮されるべき重要形質です. しかし, 植物における性決定機構はこれまで一部の植物でしか研究されておらず, 特に, 多くの動物と同様に雌雄が明確に分離する性表現(雌雄異株性)の決定機構はこれまでいずれの植物種においても明らかにされていませんでした. 本研究室では, 創設以来長きにわたって収集を続けているカキ (Diospyros kaki) とその近縁にあたるカキ属植物を用いて, その性決定機構の解明に臨んでいます.
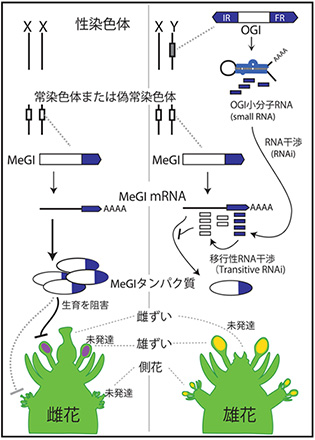
第1 図:OGI/MeGI システムによるカキ属植物の性決定メカニズム.
雄個体ではY 染色体上にあるOGI(Oppressor of meGI)が小分子RNA となって相同なMeGI(Male Growth Inhibitor)の発現を抑制する. MeGI は雄しべの発達を阻害するため, 発現量が多いと雌花になるが, OGI によって抑制されると雄花になる.
植物の性表現型は進化の中で一定ではなく, ゲノムの倍数化や栽培化の影響を受けて変化していくことが知られています. カキ属植物でも, 一般的に栽培されている「柿 (D. kaki)」では, OGI 遺伝子によって雄化が制御されているものの, かなり複雑な性表現型を示します. 今後はこのような多様性を持った性決定のメカニズム解明や, その成立・進化について考えていくとともに, キウイフルーツやブドウなどといった他科植物の性決定との共通性についても検討していきたいと思います.
参考資料
- Akagi, T., I. M. Henry, R. Tao and L. Comai. 2014. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 346: 646-650.
- Akagi, T., K. Kajita, T. Kibe, H. Morimura, T. Tsujimoto, S. Nishiyama, T. Kawai, H. Yamane, and R. Tao. 2014. Development of molecular markers associated with sexuality in Diospyros lotus L. and their application in D. kaki Thunb. J.Japan. Soc. Hort. Sci. 83: 214-221.
遺伝子組み換えと遺伝子機能評価系の開発
幼木相が長く,開花結実までに時間を要する果樹の交雑育種には長い年月と莫大な労力を要します.一方,果樹は栄養繁殖作物であるため,既存の優良品種の一部形質のみを改善すればその改良個体を接木や挿し木などの栄養繁殖により増殖してすぐにでも品種として利用することが可能です.また,最近のゲノム研究の進展により果樹の有用形質を支配する遺伝子の候補が多数,同定されてきています.このように果樹の育種や遺伝子機能評価に使える遺伝子組み換え系への需要が高まっています.しかしながら果樹の多くの種では,利用可能な効率的な遺伝子組み換えシステムが確立されていません.そこで当研究室では,ウメやオウトウなどのサクラ属果樹やカキの遺伝子組み換えシステムを確立するための研究を行っています.またアグロバクテリウム法に加えて,ウィルスベクターを用いたVIGSにより,生殖形質関連遺伝子を短期間に評価する手法の開発も試みています.

PDS遺伝子のVIGSにより葉が白化したオウトウ実生(左)と対照実生(右)
参考資料
- Kawai, T., A. Gonoi, M. Nitta, M. Kaido, N. Yamagishi, N. Yoshikawa, and R. Tao. 2014. Virus-induced gene silencing in apricot (Prunus armeniaca L.) and Japanese apricot (P. mume Siebold & Zucc.) with the Apple Latent Spherical Virus vector system. J. Japan. Soc. Hort. Sci. 83:23-31.
花成制御機構
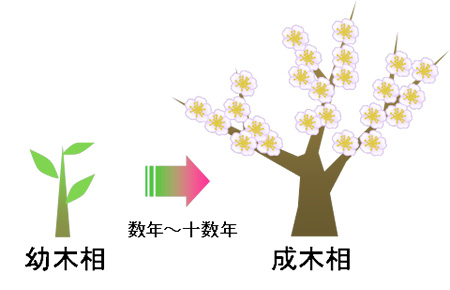
果樹の実生は,発芽後一定の年数(幼木相)を経ないと着花せず,成木相に達してはじめて安定的に花芽を着生するようになります.果樹の幼若期間は長く,交雑育種の効率化を阻む障壁です.カンキツのなかでグレープフルーツなどのブンタン類は播種後1年未満の実生が低温に遭遇するとその頂端に着花する性質(幼樹開花性)をもつことが知られています.また,ほとんどの落葉果樹が1年に1回開花する性質をもつなかで,ブルーベリーなど一部の果樹は1年に2回あるいは不時的に開花する性質をもつものがあります.これらの果樹素材を用いて果樹の花成制御機構の解明に挑んでいます.
参考資料
- Esumi, T., Y. Kitamura, C. Hagihara, H. Yamane, and R. Tao. 2010. Identification of a TFL1 ortholog in Japanese apricot (Prunus mume Sieb et Zucc.). Scientia Hort. 125: 608-616.
進化ゲノム解析
果樹作物は世代時間が非常に長く, 高度にヘテロなゲノムを有しているため交配を利用した一般的な遺伝学の適用が難しいことがあり, また, 変異体の作出も容易ではありません. そのため本研究室では, 既存の多様な種・品種を対象とし, 既に公開されている全ゲノム情報やバイオインフォマティクス技術を活用して, 主に進化学的観点からサクラ属果樹やカキ属植物において研究を始めています.
その他
「たねなし柿」として知られるカキ’平核無’(あるいは’刀根早生’)の小果枝変わり品種’突核無’の利用に関する研究や、植物ホルモンに着目した果樹の重要形質の解明などもおこなっています.

参考資料
-
Yamane, H., M. Ichiki, R. Tao, T. Esumi, K. Yonemori, T. Niikawa, and H. Motosugi. 2008. Growth characteristics of a small-fruit dwarf mutant arising from bud sport mutation in Japanese persimmon (Diospyros kaki Thunb.). HortScience 43: 1726-1730.